


























Wir verwenden Cookies, um Ihnen die optimale Nutzung unserer Webseite zu ermöglichen. Es werden für den Betrieb der Seite nur notwendige Cookies gesetzt. Details in unserer Datenschutzerklärung.
Datenschutzerklärung
Impressum
Hier beginnt der Hauptinhalt dieser Seite
Ein zentrales pflanzenbauliches, ökonomisches und umweltrelevantes Ziel im Ackerbau ist es, die in den Pflanzenaufwüchsen gespeicherten Nährstoffe im Anbausystem über mikrobielle Akkumulations- und Freisetzungsprozesse effizient zu nutzen. Dieser Kreislaufgedanke stellt in nährstofflimitierten Anbausystemen des Ökolandbaus die Grundvoraussetzung für erfolgreichen Ackerbau dar und hat im integrierten Ackerbau, im Zusammenhang mit Transformationsprozessen des Stickstoffs, seit jeher eine zentrale Bedeutung. In diesem Systemkomplex spielt der Anbau von Zwischenfrüchten eine wichtige Rolle.
Die Faktoren, die den N-Transformationsprozess aus Zwischenfrüchten in die nachfolgende Hauptfrucht beeinflussen, lassen sich wie folgt zusammenfassen:
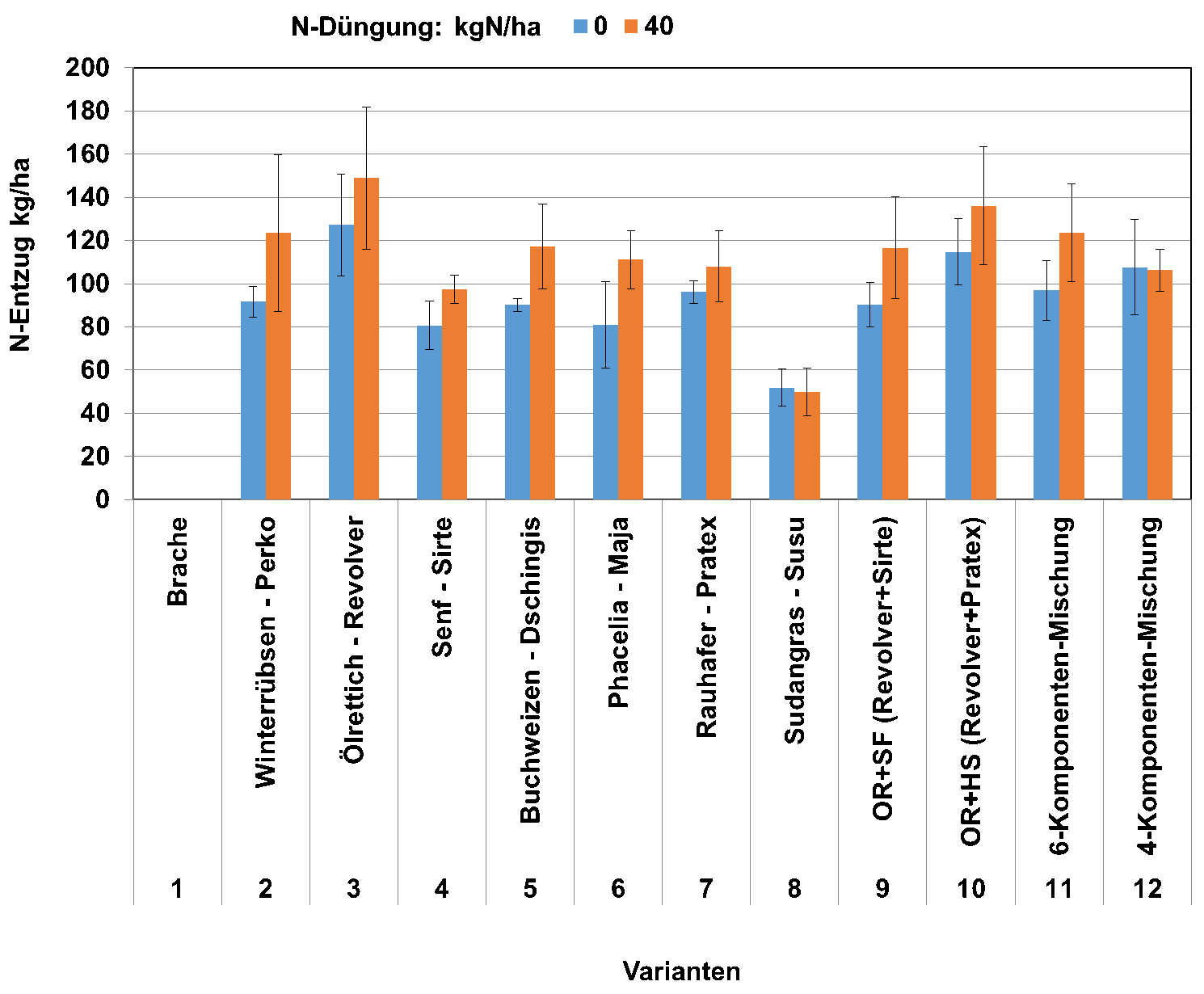
Bei frühzeitiger Saat zeigen vor allem wachstumsstarke, nichtlegume Zwischenfrüchte ein sehr hohes N-Fixierungspotenzial (Abbildung 1). Reststickstoff in Form von Nitrat kann dadurch vor der Sickerwasserperiode im Spätherbst effizient vor Auswaschung geschützt werden (Abbildung 2). Dies ist in zahlreichen Versuchen belegt.
Aus den mitunter hohen N-Mengen, die überwiegend in Proteinverbindungen insbesondere in den oberirdischen Aufwüchsen der Zwischenfrüchte zu finden sind, könnte abgeleitet werden, dass nach Mineralisierungsprozessen ein Großteil dieses Stickstoffs den nachfolgenden Kulturen wieder zur Verfügung steht. Hier zeigen sich aber sowohl in Versuchen als auch unter Praxisbedingungen sehr große Spannweiten. In der Praxis werden trotz üppiger Zwischenfruchtbestände vor Winter oftmals nur geringe Mineralisierungsleistungen im Frühjahr beobachtet.
Welche N-Mengen in welchen Zeiträumen aus der Zwischenfrucht mineralisiert werden, hängt von vielfältigen Faktoren ab, die generell Einfluss auf Mineralisierungsprozesse im Boden haben:
Entsprechend unterschiedlich können daher auch die ertragswirksamen Fruchtfolgeeffekte über den Nährstofftransfer einer Zwischenfrucht in die Hauptfrucht sein. In diesem Zusammenhang spricht man von dem Vorfruchteffekt, der in der Anbaufolge Zwischenfrucht – Hauptfrucht, oder auch Hauptfrucht nach Hauptfrucht (z.B. Winterweizen nach Raps) zu einem nicht unerheblichen Teil aus dem N-Transfer mineralisierter Pflanzenreste resultiert.
Im Rahmen der geltenden Düngeverordnung ist eine Stickstoffnachlieferung von der Zwischenfrucht in die Hauptfrucht anzurechnen. In welcher Größenordnung, hängt davon ab, ob winterharte oder abfrierende Arten beziehungsweise Leguminosen oder Nichtleguminosen als Zwischenfrüchte angebaut werden. Zudem ist zu berücksichtigen, ob Zwischenfrüchte bereits im Herbst oder erst im Frühjahr eingearbeitet werden. Daraus ergeben sich durch die Vorgaben der Düngeverordnung beziehungsweise länderspezifischen Vollzugsverordnungen Abschläge bei der Düngebedarfsermittlung der Hauptkultur. Die in der Tabelle 1 zugrunde gelegten Abschläge bei Stickstoff in der Düngebedarfsermittlung zur Hauptkultur, die einer kalkulatorischen Stickstoffnachlieferung aus der Zwischenfrucht entsprechen sollen, liegen eher im unteren Bereich. Auf besseren Standorten und bei guter Wasserversorgung kann die Stickstoffnachlieferung auch darüber liegen; bei abfrierenden Leguminosenbeständen sogar bei 60 bis 80 Kilogramm pro Hektar (Kivelitz und Stemann, 2020).
Um den zur Verfügung stehenden Stickstoff aus der Zwischenfrucht bei der Düngebedarfsermittlung (DBE) möglichst exakt anrechnen zu können, empfiehlt sich eine Nmin-Untersuchung der eigenen Flächen. Im Falle leguminosenhaltiger Mischungen, bei vorgezogenem Umbruchbeziehungsweise bei abgefrorenen Zwischenfrüchten wird der Nmin-Wert in der Regel höher sein als bei nichtlegumen, nicht eingearbeiteten beziehungsweise nicht abgestorbenen Zwischenfrüchten. Daher muss im Fall abgestorbener Zwischenfrüchte auch kein weiterer Abzug in der DBE vorgenommen werden. Die Nmin-Werte liegen auf leichten Standorten auf einem niedrigeren Niveau als auf schweren Standorten, wie die Nmin-Verlaufskurve in Abbildung 3 bei verschiedener Zwischenfrüchte veranschaulicht.
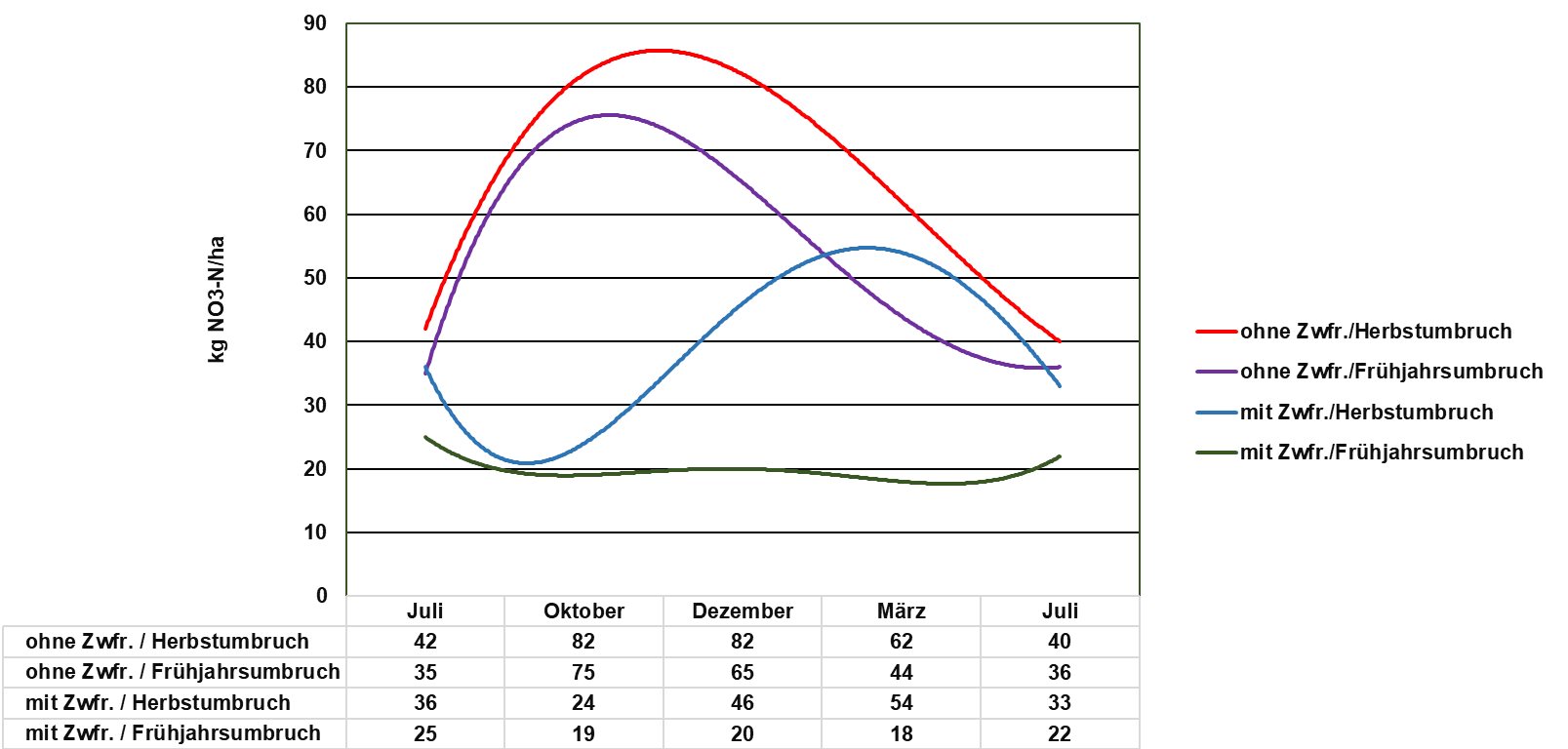
Abbildung 4 verdeutlicht auf Basis langjähriger Versuche, den typischen Verlauf des Nmin-Gehaltes in Abhängigkeit vom Umbruchtermin mit und ohne Zwischenfrucht (Herbst/Frühjahr).
Ohne Zwischenfrucht steigen die Nmin-Werte während der Sickerwasserperiode im Herbst und Winter deutlich an, unabhängig davon, ob im Frühjahr oder Herbst umgebrochen wird. Beim Herbstumbruch liegen die Nmin-Werte während der Sickerwasserperiode gegenüber dem Frühjahrsumbruch auf etwas höherem Niveau.
Solche Verlaufskurven der Mineralisierung sind aus pflanzenbaulicher Sicht und aus Gründen des Gewässerschutzes ungünstig zu bewerten, da hohe Mengen an mineralisiertem Stickstoff während der Sickerwasserperiode potenziell auswaschungsgefährdet sind und dem Pflanzenwachstum unter Umständen nicht zur Verfügung stehen. Dies gilt besonders für leichte Standorte.
Bei dem Verfahren „Zwischenfrucht mit Frühjahrsumbruch“ bleiben die Nmin-Gehalte während der gesamten Vegetationsruhe (auch während der Sickerwasserperiode) auf niedrigem Niveau. Erst nach dem Frühjahrsumbruch ist ein leichter Anstieg bis in den Juli hinein erkennbar.
Ob dieser leichte Anstieg primär aus N-Umsetzungsprozessen aus der Zwischenfrucht resultiert, kann auf Basis der zugrundeliegenden Versuchsdaten nicht zweifelsfrei belegt werden. Es kann sogar davon ausgegangen werden, dass nennenswerte N-Umsetzungsprozesse aus der Zwischenfrucht bei Frühjahrsumbruch erst zu einem späteren Zeitpunkt im Sommer stattfinden. Das heißt, dass die eher kurzlebigen Hauptkulturen wie Sommergetreide, die ihren größten Stickstoffbedarf im Mai/Juni haben, nicht hinreichend von der Stickstoffnachlieferung der Zwischenfrüchte bei Frühjahrsumbruch profitieren. Dadurch kann die oft unbefriedigende Stickstoffnachlieferung aus der Zwischenfrucht erklärt werden. Allerdings profitiert die zweite Kultur nach der Zwischenfrucht von der verzögerten Nährstoffmineralisation. Hauptkulturen, die bis in den Spätsommer hinein einen Stickstoffbedarf haben (z. B. Kartoffeln, Zuckerrüben, Mais), profitieren potenziell stärker von der späten Stickstoffnachlieferung aus der Zwischenfrucht.
Bei kritischer Betrachtung scheint der Verlauf der N-Mineralisierungskurve in Abbildung 4 beim Verfahren „Frühjahrsumbruch der Zwischenfrucht“ jedoch eher untypisch zu sein. Höchste Mineralisierungsraten werden stets bei ansteigenden Temperaturen und ausreichender Bodenfeuchte in den Monaten Mai und Juni erreicht, also einer Zeitspanne, in der viele Hauptkulturen analog zum Massenwachstum einen hohen N-Bedarf haben. Dennoch können starke Jahres- und Standorteffekte auftreten. Insbesondere bei ausgeprägter Frühjahrs- und Frühsommertrockenheit finden kaum noch Umsetzungsprozesse aus der Zwischenfrucht statt. Erst mit der Bodenbearbeitung nach der Ernte der Hauptkultur kommt auch die Mineralisierung aus der Zwischenfrucht wieder in Gang.
Im Sinne des Gewässerschutzes ist eine gute N-Versorgung der Zwischenfrüchte eine wesentliche Voraussetzung, um die ihnen zugedachten Funktionen hinreichend erfüllen zu können. Dadurch bleiben die Zwischenfruchtbestände lange in der vegetativen Phase und das C/N-Verhältnis bleibt relativ eng.
Ein frühzeitiges Mulchen solcher Zwischenfruchtbestände sollte unbedingt vermieden werden. Durch das intensive Zerkleinern der Zwischenfruchtbiomasse tritt stickstoffhaltiger Pflanzensaft aus, der im Zuge der Sickerwasserbildung in tiefere Bodenschichten verlagert werden kann. Vor allem aber finden insbesondere unter milden Bedingungen bereits Umsetzungsprozesse der Pflanzenteile mit einem engen C/N-Verhältnis statt (vor allem Blätter). Durch das Kleinhäckseln der oberirdischen Pflanzenmasse vergrößert sich die Oberfläche, was den Rotteprozess durch Pilze, Bakterien, Insekten, Regenwürmer und andere beschleunigt.
Gleiches gilt im Wesentlichen für die Einarbeitung der Zwischenfrüchte im Herbst. Die Nitratgehalte steigen, wie in Abbildung 4 dargestellt, bereits im Spätherbst und Winter deutlich an. Schnell abfrierende Zwischenfrüchte mit engem C/N-Verhältnis und hier vor allem schnell abbaubare Leguminosen sollten daher nicht vor März eingearbeitet werden.
Tabelle 2: C/N-Verhältnis von Spross und Wurzeln verschiedener Zwischenfruchtarten (Erhart et al., 2020)
| Pflanzenart | C/N oberirdisch | C/N unterirdisch |
| Kreuzblütler | ||
| Gelbsenf | 16-23 | 36-51 |
| Kresse | 14-16 | 23-32 |
| Leindotter | 12-27 | 29-61 |
| Meliorationsrettich | 16-16 | 31-38 |
| Ölrettich | 13-37 | 14-48 |
| Rübse | 7-25 | 12-38 |
| Sareptasenf | 8-13 | 18-32 |
| Sommerraps | 11-20 | 21-42 |
| Korbblütler | ||
| Ramtillkraut | 17-38 | 33-67 |
| Ringelblume | 12-15 | 24-31 |
| Sonnenblume | 23-53 | 44-141 |
| Tagetes | 10-23 | 29-66 |
| Süßgräser | ||
| Grüschnittroggen | 11 | 17-24 |
| Rispenhirse | 26-37 | 45 |
| Sandhafer (Rauhafer) | 18-28 | 24-36 |
| Sudangras | 13-47 | 28-52 |
| Leguminosen | ||
| Ackerbohne | 11-16 | 13-22 |
| Felderbse | 14-15 | 12-16 |
| Sommerwicke | 11-14 | 11-15 |
| Pannonische Wicke | 10-13 | 11-13 |
| Linse | 11-15 | 11-13 |
| Alexandrinerklee | 13-19 | 14-15 |
| Perserklee | 11-15 | 12-15 |
| Sonstige | ||
| Buchweizen | 26-37 | 47-55 |
| Lein | 13-41 | 27-65 |
| Phacelia | 14-34 | 37-82 |
| Hanf | 20-27 | 58-75 |
| Mauretanische Malve | 10-18 | 25-41 |
Der beste Effekt der Einarbeitung im Frühjahr kann bei ausreichend abgetrockneten Böden sowie trockenen und „mürben“ Zwischenfruchtresten erreicht werden. Aber auch ohne frühzeitiges Mulchen oder Einarbeiten der Zwischenfrüchte im Herbst kann ein Verlust von Stickstoff insbesondere abfrierender Zwischenfrüchte nicht vermieden werden.
Wie in Tabelle 3 verdeutlicht, reduziert sich über die Wintermonate der Stickstoffgehalt im Stängel und vor allem im Blatt. Je nach Blattanteil beziehungsweise C/N-Verhältnis des Sprosses können diese Stickstoffminderungen erheblich sein. Hier unterscheiden sich die verschiedenen Zwischenfruchtarten deutlich voneinander. Die durch das Abfrieren der Zwischenfrüchte austretenden stickstoffhaltigen Zellsäfte können somit ein nicht unerhebliches Verlustpotenzial darstellen. Damit kann ein Teil des „Transferproblems“ des Stickstoffs von der Zwischenfrucht in die Hauptfrucht erklärt werden.
Tabelle 3: Trockenmasse, Stickstoffaufnahme und das Verhältnis von Kohlenstoff zu Stickstoff im Spross vor und nach Winter 2017/18 verschiedener Zwischenfruchtarten auf den Standorten Hennef und Niederkrüchten zu unterschiedlichen Zeitpunkten (am Standort Hennef erfolgte die Analyse getrennt nach Blatt und Stängel; Stumm et al. 2019).
| Zwischenfruchtart | GD | |||||||||
| Parameter | Datum | PH | GR | SH | IK | PK | WR | ÖR | ||
| Standort Hennef | ||||||||||
| TM dt/ha | Stängel | 23.11. | 21,4 | 3,8 | 20,3 | 4,8 | 7,4 | 11,1 | 22,2 | 5,9 |
| 21.03. | 20,7 | 5,7 | 17,2 | 2,4 | 18,6 | 7,3 | ||||
| Blatt | 23.11. | 8,3 | 15,0 | 15,1 | 8,5 | 6,5 | 13,1 | 10,7 | 4,5 | |
| 21.03. | 7,9 | 10,3 | 9,1 | 5,8 | 10,3 | n.s. | ||||
| N im Spross kg/ha | Stängel | 23.11. | 51,8 | 11,4 | 40,5 | 9,6 | 20,7 | 16,2 | 42,3 | 21,2 |
| 21.03. | 45,2 | 14,1 | 24,7 | 8,8 | 31,9 | 17,3 | ||||
| Blatt | 23.11. | 40,6 | 60,0 | 58,9 | 43,5 | 38,5 | 51,7 | 56,2 | 15,4 | |
| 21.03. | 32,7 | 37,2 | 20,9 | 23,2 | 43,0 | 21,1 | ||||
| C/N-Verhältnis | Stängel | 23.11. | 16,2 | 14,8 | 22,5 | 21,5 | 15,6 | 28,5 | 21,7 | 8,5 |
| 21.03. | 20,9 | 18,1 | 31,4 | 11,6 | 25,5 | 5,2 | ||||
| Blatt | 23.11. | 8,4 | 11,4 | 11,7 | 9,3 | 8,0 | 11,4 | 8,4 | 1,5 | |
| 21.03. | 9,2 | 12,7 | 17,3 | 10,1 | 9,8 | 2,4 | ||||
| Standort Niederkrüchten | ||||||||||
| TM dt/ha | 29.11. | 30,2 | 26,6 | 36,9 | 14,7 | 34,9 | 36,5 | 4,8 | ||
| 06.03. | 29,4 | 28,2 | 32,4 | 11,2 | 33,1 | 36,5 | 11,6 | |||
| N im Spross kg/ha | 29.11. | 122,3 | 95,4 | 111,9 | 61,8 | 118,6 | 134,2 | 29,2 | ||
| 06.03. | 70,8 | 72,9 | 75,3 | 12,7 | 121,6 | 97,3 | 68,5 | |||
| C/N-Verhältnis | 29.11. | 9,5 | 11,6 | 13,7 | 10,2 | 12,0 | 10,4 | 2,0 | ||
| 06.03. | 12,5 | 12,7 | 16,9 | 12,4 | 11,0 | 11,2 | 2,3 | |||
PH - Phacelia, GR - Grünroggen, SH - Sandhafer, IK - Inkarnatklee, PK - Perserklee, WR - Winterrübsen, ÖR – Ölrettich, GD - Grenzdifferenz α = 0,05, Tukey-Test, n.n. - nicht normalverteilt, n.s. - nicht signifikant, Fehlstellen in der Tabelle sind durch niedrige und damit nicht schneidbare Bestandeshöhen zu erklären.
Besonders bei schnell abfrierenden Leguminosen, wie Perserklee oder Alexandrinerklee, sowie Grobleguminosen, mit ihrem engen C/N-Verhältnis auch in den Wurzeln, ist das potenzielle N-Verlustpotenzial über die Wintermonate am größten. Je weiter das C/N-Verhältnis im gesamten Sprossteil ist, beziehungsweise je größer der relative Stängelanteil am Spross, desto geringer sind die potenziellen N-Verluste auf diesem Wege. Eine Quantifizierung dieses Verlustpfades ist nicht ohne Weiteres möglich und von den eingangs erwähnten Einflussfaktoren abhängig.
Bei der Betrachtung des C/N-Verhältnisses der Fraktionen Stängel und Blatt zeigt sich ein sehr deutlicher Unterschied. Im Mittel der von Stumm et al. (2019) untersuchten Zwischenfrüchte (Tabelle 3) liegt das C/N-Verhältnis des Stängels bei 20:1, während es bei den Blättern bei 10:1 liegt. Diesen Zusammenhang vorausgesetzt, können Zwischenfrüchte mit hohem Stängelanteil bzw. sehr weitem C/N-Verhältnis im Stängel wie Ölrettich oder Winterrübsen als „umsetzungsträge“ eingestuft werden. Arten wie Phacelia, Grünroggen und Perserklee haben dagegen auch im Stängel ein engeres C/N-Verhältnis, wodurch ein schnellerer und höherer N-Transfer in die Folgefrucht erwartet werden kann.
In Zwischenfruchtmischungen hat vor allem der Anteil der Leguminosen maßgeblichen Einfluss auf das C/N-Verhältnis des Pflanzenbestandes und damit auf die Umsetzungsraten und -geschwindigkeiten von Stickstoff. Die C/N-Verhältnisse und die damit zusammenhängenden N-Umsetzungsraten können jedoch in Abhängigkeit vom Saattermin und der N-Versorgung deutlich variieren.
Bei früh gesäten nichtlegumen Zwischenfrüchten oder schlecht mit Stickstoff versorgten Zwischenfruchtbeständen kann ein weites C/N-Verhältnis, bezogen auf den Gesamtspross, von > 20:1 erreicht werden. Grund dafür ist, dass unter diesen Bedingungen die generative Phase früher einsetzt und damit auch die Lignifizierung des Sprosses. Bereits leichte Fröste können zu frühzeitigen Blattverlusten führen, wodurch der relative Anteil des Stängels zunimmt. Dadurch verzögert sich nach dem Winter der mikrobielle Abbau des Sprossteils und es kommt zu einem verzögerten bzw. zu einem reduzierten N-Transfer in die nachfolgende Hauptfrucht.
Durch die Aufweitung des C/N-Verhältnisses kann es zu trägen Mineralisierungsprozessen der Zwischenfrucht kommen bis hin zu einer Immobilisierung beziehungsweise Festlegung von Stickstoff. Wie Untersuchungen von Lütke Entrup et al. (1998) zur N-Dynamik bei Grasuntersaaten (Welsches Weidelgras) in Ackerbohnen und Mais belegen, fielen bei Umbruch der Grasuntersaat im Herbst die Ertragsminderungen beim nachfolgenden Winterweizen nur gering aus (-2 bis -3 dt/ha), während mit dem Frühjahrsumbruch und nachfolgendem Körnermaisanbau sehr deutliche Ertragsverluste einhergingen (-20 dt/ha).
Auch in der zweiten Folgefrucht (Weizen nach Mais) konnten noch leichte Mindererträge festgestellt werden. Erst in der dritten Folgefrucht nach dem Anbau von Welschem Weidelgras kam es zu positiven Ertragseffekten. Dies verdeutlicht die langen Zeiträume der Umsetzungsprozesse bei Zwischenfrüchten mit sehr weitem C/N-Verhältnis, wenn die Wurzelbiomasse mit einbezogen wird.
Aufgrund ihres sehr weiten C/N-Verhältnisses beziehungsweise ihres hohen Kohlenstoffanteils tragen die Wurzeln der Zwischenfrüchte primär zum Aufbau stabiler Humusformen bei. Der in der Wurzel über Proteinverbindungen gespeicherte Stickstoff kann daher, auch bezogen auf die absolute Menge, als Beitrag für die Stickstoffnachlieferung innerhalb einer Fruchtfolge vernachlässigt werden.
Eine Ausnahme bilden die Wurzeln von Leguminosen. Diese weisen ein enges C/N-Verhältnis von durchschnittlich 14 auf. Dadurch können auch die Leguminosenwurzeln durch rasche Umsetzungsprozesse potenziell wesentlich zu Stickstoffversorgung der Nachfrucht beitragen.
Ein weiterer Verlustpfad von Stickstoff, vor allem aus der abgefrorenen Zwischenfrucht, besteht in der Ausgasung von Lachgas. Lachgas ist die umgangssprachliche Bezeichnung für Distickstoffmonoxid – kurz N2O.
Es entsteht vor allem dann, wenn Mikroorganismen im Boden oder im Wasser Stickstoffverbindungen abbauen. Die Zusammenhänge, die dieser Verlustquelle zugrunde liegen und zur Quantifizierung des Verlustpotenzials, wurden in dem bundesweit angelegten Forschungsprojekt „Minderung von Treibhausgasemissionen in der Pflanzenproduktion durch standortangepasst optimierte Zwischenfruchtanbausysteme“ (THG-ZwiFru) von 2018 bis 2022 untersucht. Die faktischen N-Verluste in Kilogramm pro Hektar über Lachgas, liegen den Untersuchungen zufolge aber im niedrigen einstelligen Bereich. Allerdings hat Lachgas eine 265-mal stärkere Klimawirksamkeit als CO2.
Wir verwenden Cookies, um Ihnen die optimale Nutzung unserer Webseite zu ermöglichen. Es werden für den Betrieb der Seite nur notwendige Cookies gesetzt. Details in unserer Datenschutzerklärung.
Datenschutzerklärung
Impressum
